 2018, Vol. 25
2018, Vol. 25


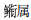 (Callionymus sp.)等。优势种随时间推移表现出较大的更替性。种类数以20 m以浅水域最多,不同水深范围的月变化有一定的差异性。20 m以浅海域的种类数与表层水温的关系在4—6月为显著正相关(r=0.997,P < 0.05),6月到7月表现为相反的变化趋势;各月的丰度与表层盐度呈显著负相关(r=-0.91,P < 0.05)。从春季到夏季,仔稚鱼高丰度区从浅水区向深水区转移。
(Callionymus sp.)等。优势种随时间推移表现出较大的更替性。种类数以20 m以浅水域最多,不同水深范围的月变化有一定的差异性。20 m以浅海域的种类数与表层水温的关系在4—6月为显著正相关(r=0.997,P < 0.05),6月到7月表现为相反的变化趋势;各月的丰度与表层盐度呈显著负相关(r=-0.91,P < 0.05)。从春季到夏季,仔稚鱼高丰度区从浅水区向深水区转移。长江口邻近水域主要指长江口门外至机轮拖网禁渔区线的河口水域。从春季至夏季, 随着长江冲淡水逐渐增强, 同时由于气温的回升, 长江口邻近水域具有一个升温降盐的过程; 到夏季以后, 由于多种海流水团的共同作用, 维持高温低盐的属性。河口水域的这种温盐度变化过程有利于海洋鱼类的产卵繁殖和饵料浮游动物的生长。因此, 长江口邻近水域是多种重要经济鱼类的产卵、育幼和索饵场[1]。东黄海鱼类大多在水温较高的春季和夏季繁殖, 因此该时间段也是河口水域仔稚鱼种类数和密度最高的季节[2-3]。长江口邻近水域繁育的经济幼鱼对于东黄海鱼类群落具有较为关键的补充作用, 也对秋冬季的渔业生产具有重要的支撑作用[4-5]。
近几十年来, 科研工作者已经对长江口水域产卵场的鱼卵仔鱼种类组成、数量分布、丰度、多样性等[6-18]进行过多次的调查研究, 这些研究成果为制定各种渔业管理和保护措施提供了有利的参考依据。随着经济的发展, 河口及近岸水域由于陆源污染物聚集、工程建设、围海造地、过度捕捞等[19-20]对经济鱼类的产卵繁殖和渔业资源补充产生了诸多不利影响。综合李建生等[21]、农牧渔业部水产局等[22]、郑元甲等[23]、程家骅等[24]、金显仕等[25]和李显森等[26]的研究结果可以看出, 与20世纪60—70年代东海渔业资源开发初期相比, 目前东海区的渔业资源量、鱼类生物学参数、群落结构和生物多样性, 均已经发生了较明显的变化; 同时, 经济种类的产卵策略、产卵时间和产卵场中心位置等可能也已发生变化。仔稚鱼阶段是鱼类生命周期中最为脆弱的时期, 其成活率的高低和存活数量是鱼类资源补充和渔业资源可持续利用的基础。因此, 有必要通过系统的调查研究来重新确定河口邻近水域仔稚鱼优势种组成、数量结构及其与环境的关系, 并以此来评估该产卵场主要经济鱼类的繁殖特征及其对近海渔业的资源补充能力。以往大多数研究者对河口水域的仔稚鱼调查研究主要以个别月份调查为主, 站位的经向设置间隔以10'或15'居多, 个别研究甚至间隔30'。李建生等[12]和刘淑德等[13]的研究结果表明, 长江口水域, 从近岸向外海, 鱼卵仔鱼的种类数和丰度均呈现降低的趋势, 并且在123°00'E以东的海域明显偏低。因此, 在近岸水域进行经向密集设站调查, 有利于对该水域鱼卵仔鱼进行系统研究。本研究拟通过春夏季在长江口邻近水域的逐月连续高密度定点调查数据, 研究其仔稚鱼的种群结构、优势种组成和丰度的月变化特征, 探讨它们的早期生态特性及与环境的关系, 为今后进一步研究东海区重要产卵场渔业资源的补充机制、渔业资源的生态修复、制订资源养护管理与可持续发展措施提供科学依据和技术支撑。
1 材料与方法 1.1 调查时间和方法调查时间为2015年4月至7月的每月中旬, 在长江口近岸的31°30'N、122°00'E~122°55'E和31°00'N、122°04'E~123°14'E分别设置两个断面, 每个断面从近岸向外海至禁渔区线, 按照经向间隔5'设置1个调查站位。两个断面分别设置12个和15个站位, 共设置27个采样站(图 1)。利用大型浮游生物网进行鱼卵和仔稚鱼采集, 大型浮游生物网具规格为口径130 cm、长280 cm、孔径0.50 mm。每站分别进行水平拖网调查, 每站拖曳10 min, 拖速2~3 kn, 起网时记录流量计(HYDRO- BIOS, 德国)数据以计算滤水体积; 采集的鱼卵和仔稚鱼样品用5%甲醛溶液固定, 在室内对样品进行种类鉴定。所有采样站位利用Seabird37型CTD进行现场温度、盐度和深度的测定。

|
图 1 长江口邻近水域调查站位 Fig.1 Sampling stations off the Yangtze River estuary |
调查水域各站位的水深范围为4~63 m (由于潮汐的变化, 同一站位在不同月份的深度会有所变化)。根据水深变化特征, 把调查站位分为3个档次, 分别是: (1) 20 m以浅水域, 共13个调查站位, 分别是N01~N06、S01~S07站; (2) 20~30 m水深, 共7个调查站位, 分别是N07~N10、S08~ S10站; (3) 30 m以深水域, 共7个调查站位, 分别是N11~N12、S11~S15站。
1.3 数据处理方法定量分析时, 各站位丰度值以仔稚鱼出现数量和流量计记录的数据换算成单位水体的仔稚鱼数量[ind/(100m3)]为指标进行相关分析。利用相对重要性指数[27](index of relative importance, IRI)对仔稚鱼重要性进行分析。由于仔稚鱼个体很小, 故分析时只采用个体数密度, 不考虑生物量密度。相对重要性指数计算公式为: IRI=(N%×F%)× 10000。其中, N%指某种类占总尾数的百分比; F%指某种类出现站位频率。仔稚鱼重要性的判断标准[15]如下: IRI大于500的为优势种, IRI在100至500之间的为重要种, IRI在10至100之间的为常见种, IRI小于10的为偶见种。把优势种、重要种和常见种都作为主要种类进行研究。利用统计软件SPSS18.0进行相关性检验分析。
2 结果与分析 2.1 种类数和丰度的月变化2015年春夏季, 在长江口邻近水域共采获35839尾仔稚鱼, 其中从4月至7月各航次分别采获1595、2348、7814、24082尾。经鉴定, 共出现62种仔稚鱼, 分别属于14目、30科、45属(表 1), 其中47个种类鉴定到种, 11个种类鉴定到属, 4个种类鉴定到科。长江口邻近水域仔稚鱼种类数和丰度的月相变化如图 2所示。由图 2可见, 分月份来看, 以6月的出现种类数最多, 为36种; 其余3个月的出现种类数从高到低分别是7月(24种)、5月(23种)、4月(22种)。随着时间的推移, 从4月到7月, 长江口邻近水域出现的仔稚鱼种类数先是缓慢增加然后快速增加, 至6月达到高峰期, 每月增加幅度分别为4.54%和56.52%;然后到7月则大幅降低, 下降幅度为33.33%。各月仔稚鱼的丰度变化范围为11.45~114.31 ind/(100 m3), 4月最低, 7月最高; 随着时间的推移, 丰度持续增加; 从4月到6月, 丰度的每月增加幅度相对较低, 分别为35.55%和61.98%, 而从6月到7月, 丰度则表现为大幅增加的趋势, 达到4个月的最高峰, 相对前3个月分别增加了8.98倍、8.63倍、7.79倍。

|
图 2 长江口邻近水域仔稚鱼种类数和丰度的月变化 Fig.2 Monthly variation in species numbers and abundance of larvae and juveniles off the Yangtze River estuary |
|
|
表 1 春夏季长江口邻近水域仔稚鱼种类组成 Tab.1 Species composition of larvae and juveniles off the Yangtze River estuary in spring and summer |
根据相对重要性指数计算结果, 长江口邻近水域仔稚鱼主要种类的月变化如表 2所示。由表 2可见, 春夏季, 该水域出现的仔稚鱼主要种类数以6月最多(13种), 5月次之(8种), 7月最少(5种)。分月份来看, 4月有3种优势种, 分别为斑尾刺虾虎鱼(Acanthogobius ommaturus)、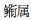
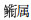
|
|
表 2 长江口邻近水域仔稚鱼主要种类相对重要性指数的月变化 Tab.2 Monthly variation in IRI of dominant larvae and juveniles off the Yangtze River estuary |
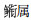 Callionymus sp.
Callionymus sp.
由不同水深范围仔稚鱼种类数的月变化结果(图 3)可见, 总体来看, 春夏季各水深范围的仔稚鱼出现种类数以20 m以浅水域最多(41种), 30 m以深水域次之(31种), 20~30 m水深最少(30种)。从各月不同水深范围仔稚鱼的种类数变化来看: 4月, 20~30 m和30 m以深海域种类数较多(均为12种), 20 m以浅海域较少; 5—6月, 均以20 m以浅水域最多, 30 m以深水域次之, 20~30 m水深最少; 7月, 以20 m以浅海域最多, 20~30 m水域次之, 30m以深水域最少。从各水深范围仔稚鱼的种类数月变化情况来看, 20 m以浅水域在4月最低, 5—6月达到高峰期, 7月又有一定程度的减少; 20~30 m水深水域, 各月种类数变化在9~12种, 月间的差异较小; 30 m以深水域, 4—5月较低, 6月迅速升高且达到高峰期, 7月又出现快速下降趋势。

|
图 3 长江口邻近水域不同水深范围仔稚鱼种类数的月变化 Fig.3 Monthly variation of species numbers of larvae and juveniles in different range of depth off the Yangtze River estuary |
由不同水深水域仔稚鱼丰度的月变化结果(图 4)可见, 4月, 20 m以浅水域的丰度最高, 20~30 m水域次之, 30 m以深水域最低; 5月, 各水深水域的仔稚鱼丰度变化趋势与4月相一致, 但20 m以浅水域仔稚鱼丰度有较大幅度的升高, 其他水深范围则略有降低; 6月, 30 m以深水域仔稚鱼的丰度最高, 20 m以浅水域次之, 20~30 m水深水域最低; 7月, 20~30 m水深水域仔稚鱼丰度最高, 30 m以深水域次之, 20 m以浅水域最低。

|
图 4 长江口邻近水域不同水深范围仔稚鱼丰度的月变化 Fig.4 Monthly variation of abundance of larvae and juveniles in different range of depth off the Yangtze River estuary |
长江口邻近水域不同水深范围平均表层水温和盐度的月变化统计结果(表 3)显示, 从4月到7月, 各个水深范围的平均表温都呈现出升高的趋势, 其中从4月到5月的升温趋势较快, 5月和6月基本持平, 6—7月又有小幅升温; 3个水深范围表层盐度的变化趋势有所差别, 20 m以浅水域为持续降低的趋势, 其他2个水深水域则是先降低后升高的趋势。为了分析不同水深水域仔稚鱼种类数和丰度与表层水温和盐度随时间变化的相关关系, 利用统计软件对其两两进行相关性检验。结果表明, 20 m以浅水域, 仔稚鱼种类数与表层水温的关系在4—6月表现为显著正相关关系(r=0.997, P < 0.05), 6月到7月表现为相反的变化趋势; 丰度与表层盐度随月份的变化表现为显著负相关关系(r=-0.91, P < 0.05)。其他2个水深水域, 仔稚鱼种类数和丰度与表层水温和盐度的月变化之间均无明显的相关性。
|
|
表 3 长江口邻近水域不同水深范围平均表层水温和盐度的月变化 Tab.3 Monthly variation in average water temperature and salinity off the Yangtze River estuary |
长江口邻近水域作为东海鱼类的主要产卵场和保育场, 其仔稚鱼群落是河口及邻近水域渔业资源补充的重要来源之一[23], 对该水域仔稚鱼的调查研究有助于今后制定更加完善的物种保护措施。本研究由于调查持续时间较长、调查次数较多、经向跨度较大, 因此仔稚鱼的种类数多达62种。相对于杨东莱等[7]20世纪80年代春夏季在长江口大范围海域连续逐月调查得到的85种鱼卵仔鱼相比, 本次调查的种类数有一定幅度的下降。但相对于20世纪90年代以来的数次单月或不连续月份调查[8-18], 则本次调查的种类数更为丰富, 也能够更真实地反映出春夏季长江口邻近水域的仔稚鱼组成和变化特征。春夏季是海洋鱼类的主要产卵时间, 在东黄海近岸水域, 随着水温从南到北逐步回升, 鱼类的产卵时间也表现出南早北晚的规律, 长江口水域鱼类的产卵高峰期为4—6月[22]。从各月调查结果来看, 仔稚鱼种类数以6月最多, 其他月份明显较低。随着水温的回升, 长江口邻近水域的鱼类主要在春末的4—5月集中产卵[7-17], 而鱼卵孵化到仔稚鱼的时间, 各种鱼类从几小时到几十天不等[28], 因此集中表现为6月的仔稚鱼种类数高达36种。上述结果表明, 长江口水域仍然是东海近海和河口水域主要及次要经济鱼类如小黄鱼、鳀、龙头鱼、虾虎鱼类、小公鱼类、凤鲚等的主要产卵和育幼场[1, 22-23]。从4月到7月, 仔稚鱼的丰度表现出前期(4—6月)缓慢增加而后期(7月)大幅增加的趋势, 与种类数的变化有较大差异。通过对调查数据的统计发现, 7月的小公鱼属、红狼牙虾虎鱼和黄姑鱼等3个种类的丰度明显偏高, 它们的累计丰度占总丰度的百分比高达87.83%, 但它们的出现率却均低于35%。因此, 可以判断后期仔稚鱼丰度大幅增加是由于少数种类的丰度在局部海域大幅提高引起的。由此推测产生上述丰度月变化的原因可能为, 海洋鱼类的产卵时间主要受水温的调控, 春末夏初, 随着海洋水温上升到一定的阈值, 河口水域的各种鱼类都逐渐开始产卵。仔稚鱼丰度的高低主要与鱼卵的孵化率以及仔稚鱼的成活率有关[23]。前期由于水温较低, 导致鱼卵的孵化成功率和仔稚鱼的饵料生物丰度都相对较低, 上述原因也使孵化成功的仔稚鱼死亡率较高, 因此丰度较低; 后期随着水温的升高, 鱼卵的孵化成功率和仔稚鱼的饵料生物丰度都相应地有所升高, 因此仔稚鱼的丰度也表现出大幅升高的趋势。
3.2 长江口邻近水域仔稚鱼优势种类的月变化特征分析本研究表明, 各月的优势种表现出较大的更替性。斑尾刺虾虎鱼和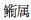
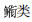
随着环境条件的变化, 仔稚鱼种类数和丰度也会表现出相应的变化规律, 但不同月份之间由于冲淡水、海流、季风等因素的变化也会有差异[23]。本研究表明, 在长江口邻近水域, 各月份仔稚鱼的种类数总体上以20 m以浅水域最多, 其他水域明显减少, 从里向外表现为降低的趋势; 而20 m以浅水域仔稚鱼种类数与表层水温的关系在4—6月表现为显著正相关关系。说明春夏季长江口近岸浅水海域是主要的鱼类产卵场, 产卵鱼类随着时间的变化是逐渐增多的, 并且在5—6月达到高峰期。不同水深范围之间, 仔稚鱼种类数的月间变化具有一定的差异性。造成差异的原因可能为, 调查海域鱼类的生态类型主要有河口型、近岸型、近海型[15], 不同类型鱼类产卵时的温度、盐度、透明度和水深都具有一定的差异性; 长江口水域具有错综复杂的海洋环境, 受各项水文因子变化所制约, 不同水深范围的鱼类种类组成和数量分布具有季节变化。各水深范围仔稚鱼丰度的变化与种类数有较大的差异。本文的研究结果显示, 春季(4—5月)的丰度以20 m以浅水域最高, 但是到夏季以后上述趋势发生了较大的变化, 20 m以浅水域的仔稚鱼丰度自6月开始低于30 m以深水域, 至7月已经远低于其他2个水深水域; 20 m以浅水域的仔稚鱼丰度与盐度随时间的推移呈现显著负相关关系。由此表明, 从春季到夏季, 仔稚鱼高丰度区从浅水区向深水区转移; 由于冲淡水的逐渐增强而形成的水温升高和盐度降低所营造的高温低盐环境, 更有利于仔稚鱼早期的生长发育及索饵。高丰度区转移原因可能有以下三点: (1)不同月份产卵场中心位置的变化。由于水温、盐度、场势的季节消长等环境因素[10]的变化, 产卵亲鱼本身具有一定的适应性变化, 导致不同月份之间的中心产卵场具有一定的变化。这种亲鱼的行为特征可直接影响到仔稚鱼丰度的空间分布。(2)仔稚鱼在近海与河口间被动漂移[23]。春夏季, 中国东海主要受西南季风的影响[22]; 同时, 该时间段的长江径流也是一个从枯水到丰水的过程, 特别是5月以后, 中国南方进入雨季, 长江径流量的增长是比较明显的。在风场和流场的共同作用下, 仔稚鱼被动地从近岸向外海漂移。(3)仔稚鱼自身的游泳行为。仔稚鱼发育到后期阶段, 已经具备一定的游泳能力, 因此会随长江径流量的多少、索饵水域的盐度和透明度的变化而表现出趋近岸或离岸的移动行为[7]。
| [1] |
Luo B Z. Ecological characteristics and fishery resources of the Yangtze river estuary and adjacent sea[J]. Resources and environment in the Yangtze Basin, 1992, 1(1): 24-30. [罗秉征. 河口及近海的生态特点和渔业资源[J]. 长江流域资源和环境, 1992, 1(1): 24-30.] |
| [2] |
Barletta-Bergan A, Barletta M, Salint-Paul U. Structure and seasonal dynamics of larval fish in the Caeté River Estuary in North Brazil[J]. Estuarine, Coastal and Shelf Science, 2002, 54(2): 193-206. DOI:10.1006/ecss.2001.0842 |
| [3] |
Liu S D, Xian W W, Liu D. Characteristics of ichthyoplankton assemblages in Yangtze Estuary and adjacent waters in spring[J]. Chinese Journal of Applied Ecology, 2008, 19(10): 2284-2292. [刘淑德, 线薇薇, 刘栋. 春季长江口及邻近海域鱼类浮游生物群落特征[J]. 应用生态学报, 2008, 19(10): 2284-2292.] |
| [4] |
Li J S, Li S F, Ren Y P, et al. Seasonal variety of fishery biology community structure in fishing ground of the Yangtze estuary[J]. Journal of Fishery Sciences of China, 2004, 11(5): 432-439. [李建生, 李圣法, 任一平, 等. 长江口渔场渔业生物群落结构的季节变化[J]. 中国水产科学, 2004, 11(5): 432-439. DOI:10.3321/j.issn:1005-8737.2004.05.009] |
| [5] |
Cheng J H, Zhang Q H, Li S F, et al. Utilizing to the Fisheries Resources in the East China Sea and Yellow Sea[M]. Shanghai: Shanghai Scientific & Technical Publishers, 2006: 1-22. [程家骅, 张秋华, 李圣法, 等. 东黄海渔业资源利用[M]. 上海: 上海科学技术出版社, 2006: 1-22.]
|
| [6] |
Wu G Z. The ecological characteristics of distribution of eggs, larvae and juveniles of the Engraulis japonicus (Temminck & Schlegel) and Anchoviella commersonii (Lecépède) in the Changjiang River estuary[J]. Oceanologia et Limnologia Sinica, 1989, 20(3): 217-229. [吴光宗. 长江口海区鳀鱼和康氏小公鱼鱼卵和仔、稚鱼分布的生态特征[J]. 海洋与湖沼, 1989, 20(3): 217-229.] |
| [7] |
Yang D L, Wu G Z, Sun J R. The investigation of pelagic eggs, larvae and juveniles of fishes at the mouth the Changjiang River and adjacent areas[J]. Oceanologia et Limnologia Sinica, 1990, 21(4): 346-355. [杨东莱, 吴光宗, 孙继仁. 长江口及其邻近海区的浮性鱼卵和仔稚鱼的生态研究[J]. 海洋与湖沼, 1990, 21(4): 346-355.] |
| [8] |
Xu Z L, Yuan Q, Jiang M, et al. An investigation of fish eggs, larvae and juveniles in the Changjiang estuary[J]. Journal of Fishery Sciences of China, 1999, 6(5): 63-64. [徐兆礼, 袁骐, 蒋玫, 等. 长江口鱼卵和仔、稚鱼的初步调查[J]. 中国水产科学, 1999, 6(5): 63-64.] |
| [9] |
Zhu X H, Liu D, Sha X S. Relationship between community structure of ichthyoplankton and environmental factors in the estuary of Changjiang River in spring[J]. Studia Marina Sinica, 2002, 44: 169-179. [朱鑫华, 刘栋, 沙学绅. 长江口春季鱼类浮游生物群落结构与环境因子的关系[J]. 海洋科学集刊, 2002, 44: 169-179.] |
| [10] |
Shan X J, Xian W W, Wu Y F. Dynamic changes in the ichthyoplankton community structure after the sluice of the Three Gorges Dam[J]. Periodical of Ocean University of China, 2005, 35(6): 936-940. [单秀娟, 线薇薇, 武云飞. 三峡工程蓄水前后秋季长江口鱼类浮游生物群落结构的动态变化初探[J]. 中国海洋大学学报(自然科学版), 2005, 35(6): 936-940.] |
| [11] |
Jiang M, Shen X Q, Wang Y L, et al. Species of fish eggs and larvae and distribution in Changjiang Estuary and vicinity waters[J]. Acta Oceanologica Sinica, 2006, 28(2): 171-174. [蒋玫, 沈新强, 王云龙, 等. 长江口及其邻近水域鱼卵、仔鱼的种项组成与分布特征[J]. 海洋学报(中文版), 2006, 28(2): 171-174. DOI:10.3321/j.issn:0253-4193.2006.02.023] |
| [12] |
Li J S, Hu F, Cheng J H. Distributions of fish eggs and juveniles and relations between surface layer water temperature and salinity of the Yangtze estuary water area in spring[J]. Marine Sciences, 2007, 31(4): 16-21. [李建生, 胡芬, 程家骅. 长江口水域春季鱼卵仔鱼分布及其与温度盐度的关系[J]. 海洋科学, 2007, 31(4): 16-21. DOI:10.3969/j.issn.1000-3096.2007.04.005] |
| [13] |
Liu S D, Xian W W. Temporal and spatial patterns of the ichthyoplankton community in the Yangtze Estuary and its adjacent waters[J]. Biodiversity Science, 2009, 17(2): 151-159. [刘淑德, 线薇薇. 长江口及其邻近水域鱼类浮游生物群落的时空格局[J]. 生物多样性, 2009, 17(2): 151-159.] |
| [14] |
Wan R J, Zhou F, Shan X J, et al. Impacts of variability of habitat factors on species composition of ichthyoplankton and distribution of fish spawning ground in the Changjiang River estuary and its adjacent waters[J]. Acta Ecologica Sinica, 2010, 30(3): 155-165. DOI:10.1016/j.chnaes.2010.04.006 |
| [15] |
Li J S, Hu F, Lin N. Ecological distribution of fish larvae and juveniles in the Yangtze River estuary and its adjacent waters in spring[J]. South China Fisheries Science, 2015, 11(1): 1-8. [李建生, 胡芬, 林楠. 长江口及邻近海域春季仔、稚鱼的生态分布研究[J]. 南方水产科学, 2015, 11(1): 1-8. DOI:10.3969/j.issn.2095-0780.2015.01.001] |
| [16] |
Yu H C, Xian W W. The environment effect on fish assemblage structure in waters adjacent to the Changjiang (Yangtze) River estuary (1998-2001)[J]. Chinese Journal of Oceanology and Limnology, 2009, 27(3): 443-456. DOI:10.1007/s00343-009-9155-6 |
| [17] |
Liu Y Y, Song C, Zhao F, et al. Category composition and distributional patterns of fish larvae and juveniles in the Yangtze Estuary and adjacent waters[J]. Marine Fisheries, 2016, 38(6): 597-608. [刘媛媛, 宋超, 赵峰, 等. 长江口及其邻近水域仔稚鱼种类组成及分布特征[J]. 海洋渔业, 2016, 38(6): 597-608. DOI:10.3969/j.issn.1004-2490.2016.06.005] |
| [18] |
Jiang M, Shen X Q, LI L, et al. Annual variability of ichthyoplankton in the Yangtze River estuary of China from August 2002 to 2009[J]. Oceanological and Hydrobiological Studies, 2013, 42(1): 59-69. |
| [19] |
Zhang Q H, Cheng J H, Xu H X, et al. The Fishery Resources and Its Sustainable Utilization in the East China Sea Region[M]. Shanghai: Fudan University Press, 2007: 121-133. [张秋华, 程家骅, 徐汉祥, 等. 东海区渔业资源及其可持续利用[M]. 上海: 复旦大学出版社, 2007: 121-133.]
|
| [20] |
Dong F Y. A preliminary prediction on the effects of the eastern route of water transferring project from Yangtze River on fisheries resources in Yangtze River estuary[J]. Resources and Environment in the Yangtze Valley, 1997, 6(2): 168-172. [董方勇. 南水北调东线工程对长江口渔业资源的影响[J]. 长江流域资源与环境, 1997, 6(2): 168-172.] |
| [21] |
Li J S, Li S F, Ding F Y, et al. Analysis on annual change of fish diversity in Yangtze estuary offshore water area[J]. Journal of Fishery Sciences of China, 2007, 14(4): 637-643. [李建生, 李圣法, 丁峰元, 等. 长江口近海鱼类多样性的年际变化[J]. 中国水产科学, 2007, 14(4): 637-643. DOI:10.3321/j.issn:1005-8737.2007.04.016] |
| [22] |
Fishery Bureau of Ministry of Agriculture, Command Post of East China Sea Fishery of Ministry of Agriculture. Investigation and Area Division of Fisheries Resources in the East China Sea Region[M]. Shanghai: East China Normal University Press, 1987: 216-622. [农牧渔业部水产局, 农牧渔业部东海区渔业指挥部. 东海区渔业资源调查和区划[M]. 上海: 华东师范大学出版社, 1987: 216-622.]
|
| [23] |
Zheng Y J, Chen X Z, Cheng J H, et al. The Fisheries Resources and Environment in the East China Sea Shelf[M]. Shanghai: Shanghai Scientific & Technical Publishers, 2003: 206-741. [郑元甲, 陈雪忠, 程家骅, 等. 东海大陆架生物资源与环境[M]. 上海: 上海科学技术出版社, 2003: 206-741.]
|
| [24] |
Cheng J H, Ding F Y, Li S F, et al. Changes of fish community structure in the coastal zone of the northern part of East China Sea[J]. Journal of Natural Resources, 2006, 21(5): 775-781. [程家骅, 丁峰元, 李圣法, 等. 夏季东海北部鱼类群落结构的变化[J]. 自然资源学报, 2006, 21(5): 775-781. DOI:10.3321/j.issn:1000-3037.2006.05.011] |
| [25] |
Jin X S, Shan X J, Guo X W, et al. Community structure of fishery biology in the Yangtze River estuary and its adjacent waters[J]. Acta Ecologica Sinica, 2009, 29(9): 4761-4772. [金显仕, 单秀娟, 郭学武, 等. 长江口及其邻近海域渔业生物的群落结构特征[J]. 生态学报, 2009, 29(9): 4761-4772. DOI:10.3321/j.issn:1000-0933.2009.09.021] |
| [26] |
Li X S, Yu Z H, Sun S, et al. Ecological niche breadth and nicheoverlap of dominant species of fish assemblage in Yangtze River estuary and its adjacent waters[J]. Chinese Journal of Applied Ecology, 2013, 24(8): 2353-2359. [李显森, 于振海, 孙珊, 等. 长江口及其毗邻海域鱼类群落优势种的生态位宽度与重叠[J]. 应用生态学报, 2013, 24(8): 2353-2359.] |
| [27] |
Pinkas L, Oliphant M S, Iverson I L K. Food habits of albacore, bluefin tuna, and bonito in California waters[J]. Fish Bulletin, 1971, 152: 1-105. |
| [28] |
Zhao C Y, Zhang R Z. The Fish Eggs and Larvae in Coastal Waters of China[M]. Shanghai: Shanghai Scientific & Technical Publishers, 1985: 9-12. [赵传絪, 张仁斋. 中国近海鱼卵与仔鱼[M]. 上海: 上海科学技术出版社, 1985: 9-12.]
|
| [29] |
Zhuang P, Wang Y H, Li S F, et al. Fishes of the Yangtze Estuary[M]. Shanghai: Shanghai Scientific & Technical Publishers, 2006: 207-223. [庄平, 王幼槐, 李圣法, 等. 长江口鱼类[M]. 上海: 上海科学技术出版社, 2006: 207-223.]
|