 2020, Vol. 27
2020, Vol. 27


2. 中国水产科学研究院长江水产研究所, 湖北 武汉 430223;
3. 华中农业大学水产学院, 池塘健康养殖湖北省工程实验室, 湖北 武汉 430070;
4. 贵州大学动物科学学院, 贵州 贵阳 550025;
5. 贵州大学, 高原山地动物遗传育种与繁殖教育部重点实验室, 贵州 贵阳 550025
2. Yangtze River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Wuhan 430223, China;
3. College of Fisheries, Hubei Provincial Engineering Laboratory for Pond Aquaculture, Huazhong Agricultural Universtiy, Wuhan 430070, China;
4. College of Animal Science, Guizhou University, Guiyang 550025, China;
5. Key Laboratory of Animal Genetics, Breeding and Reproduction in the Plateau Mountainous Region, Ministry of Education; Guizhou University, Guiyang 550025, China
受人类活动的干扰, 许多鱼类资源已严重衰竭, 一些种类甚至已经逼近灭绝的边缘。鱼类是水域生态系统的重要组成部分, 其资源量的变动、消失或者迁徙将通过生态级联效应引起水域生态系统结构和功能的失衡, 最终给水生生物多样性和生态平衡带来严重威胁。为此, 大量的养护措施, 例如, 最小起捕个体、禁渔期、禁渔区以及过鱼通道等, 被不加区别地广泛地用于鱼类资源的养护, 然而, 所实施养护措施的科学性和合理性往往受到忽视[1-3]。
异齿裂腹鱼(Schizothorax oconnori Lloyd)是栖息于青藏高原的一种鲤科冷水性鱼类, 是该地区重要特色鱼类, 在研究生物地理学和鱼类演化等方面具有极为重要的科学参考价值。在特殊而恶劣的雅鲁藏布江水域生态环境的长期选择下, 异齿裂腹鱼形成了种群高度单一和一致、年龄结构复杂、初次产卵时间晚、怀卵量低以及生长缓慢等生物学特征, 上述特征使其对人类的生产活动, 尤其是捕捞活动的响应十分敏感[4-8]。近十几年来, 随着人类活动加剧和西藏地区生态环境改变, 以及受过度捕捞和外来物种入侵等因素影响, 雅鲁藏布江流域异齿裂腹鱼自然种群数量急剧减少[9-13], 对其现存种群的资源养护工作亟待开展。然而, 目前已有的报道绝大部分集中于对异齿裂腹鱼资源现状的定性描述[6, 9-10], 定量描述异齿裂腹鱼所占渔获物的比重、种群资源状况以及评估不同养护措施对其资源保护效果的研究仍然稀少, 仅见杨汉运等[11]通过对雅鲁藏布江中游5个江段的渔获物调查, 该研究发现异齿裂腹鱼在雅鲁藏布江中游分布较为均匀, 是整个中游江段的优势种群, 占中游渔获物总重的37.60%。相关研究的匮乏严重制约了异齿裂腹鱼资源养护工作的开展。
本研究利用单位补充量模型评估了雅鲁藏布江流域异齿裂腹鱼的种群资源现状, 并分析了14种渔业管理政策对异齿裂腹鱼种群资源的养护效果, 以期为科学合理地开展其资源养护工作提供基础数据。
1 材料和方法 1.1 样本采集2008年8月至2009年8月在雅鲁藏布江拉孜至尼木江段使用刺网和地笼等渔具逐月采集异齿裂腹鱼样本(图 1)。每月采集样本不少于30尾, 共采集样本1126尾。测量体长(standard length, SL)和体重(body weight, W), 长度精确到1 mm, 重量精确到0.1 g。解剖后, 取一对微耳石用于年龄鉴定, 并肉眼辨别性别。

|
图 1 雅鲁藏布江中游异齿裂腹鱼采样点 Fig.1 Sampling locations in the middle reaches of Yarlung Zangbo River for Schizothorax oconnori |
估算的异齿裂腹鱼的von Bertalanffy体长生长方程如下[6]:
| $ 雌性:{L_t} = 576.9\left[ {1 - {{\rm{e}}^{ - 0.081(t + 0.946)}}} \right] $ | (1) |
| $ 雄性:{L_t} = 499.7\left[ {1 - {{\rm{e}}^{ - 0.095(t + 0.896)}}} \right] $ | (2) |
式中, Lt为异齿裂腹鱼在t龄时的体长。
利用异齿裂腹鱼的体长−体重公式[6], 将其体长数据转换成体重数据, 体长–体重公式如下:
| $ 雌性:W = {\rm{ }}8.897 \times {10^{-6}}{\rm{S}}{{\rm{L}}^{3.080}} $ | (3) |
| $ 雄性:W = {\rm{ }}8.327 \times {10^{-6}}{\rm{S}}{{\rm{L}}^{3.090}} $ | (4) |
异齿裂腹鱼每年产卵1次, 是典型的春季产卵鱼类, 产卵季节为3—5月[6]。异齿裂腹鱼在t龄时其种群中性成熟个体所占比例采用Logistic方程来估算(未发表数据), 具体计算公式如下:
| $ 雌性:{G_t} = {\rm{ }}1{\rm{ }}/\left\{ {1{\rm{ }} + {\rm{ exp }}\left[ { - 0.472{\rm{ }}\left( {A - 9.5} \right)} \right]} \right\} $ | (5) |
| $ 雄性:{G_t} = {\rm{ }}1{\rm{ }}/\left\{ {1{\rm{ }} + {\rm{ exp }}\left[ { - 0.261\left( {A - 7.2} \right)} \right]} \right\} $ | (6) |
总死亡率, 采用Chapman-Robson法估算总死亡率Z [14], 公式如下:
| $ \begin{array}{l} Z = \ln \frac{{1 + \bar T - {t_{\rm{c}}} - \frac{1}{N}}}{{\bar T - {t_{\rm{c}}}}} - \\ \;\;\;\;\frac{{\left( {N - 1} \right)\left( {N - 2} \right)}}{{N\left[ {N\left( {\bar T - {t_{\rm{c}}}} \right) + 1} \right]\left[ {N + N\left( {\bar T - {t_{\rm{c}}}} \right) - 1} \right]}} \end{array} $ | (7) |
式中, tc为起捕年龄; T为样本中年龄不小于tc的所有个体的平均年龄; N为样本中年龄不小于tc的个体的数量。
自然死亡率, 雌鱼和雄鱼的自然死亡率(M)分别使用以下两个经验方程式来计算:
| $ M = 4.118{k^{0.73}}L_\infty ^{ - 0.33\;[15]} $ | (8) |
| $ M = {\rm{ }} - {\rm{ln}}\left( {0.01} \right)/{t_{{\rm{max}}}}^{[16]} $ | (9) |
式中, L∞为渐进体长(单位为cm); k为生长系数; tmax为最大年龄, 计算公式为: tmax = t0+ 2.996/k, 其中k为生长系数, t0为体长为0时的年龄[17]。
捕捞死亡率, 当前的捕捞死亡率Fcur = Z-M。假设估算的捕捞死亡率和自然死亡率在不同龄组间保持恒定。
1.5 种群动态评析 1.5.1 评估模型采用单位补充量模型和繁殖潜力比评估异齿裂腹鱼的资源状况及其对不同渔业养护措施的响应[18-19], 模型输入参数见表 1。对传统的单位补充量模型进行修改, 即将时间递增单位修改为月, 以便于评估禁渔期对繁殖潜力比(SPR)和单位补充量渔获量(YPR)的影响。
|
|
表 1 异齿裂腹鱼单位补充量模型输入参数 Tab.1 The input parameters for per-recruit models of Schizothorax oconnori |

|
图 2 雅鲁藏布江异齿裂腹鱼捕捞曲线 Fig.2 Catch curve of Schizothorax oconnori in the Yarlung Zangbo River |
(1) 繁殖潜力比(SPR)计算公式如下:
| $ {\rm{SPR}} = \frac{{{\rm{SSB}}{{\rm{R}}_F}}}{{{\rm{SSB}}{{\rm{R}}_{F = 0}}}} $ | (10) |
| $ \begin{array}{l} {\rm{SSBR}} = \frac{{{\rm{SSB}}}}{R} = \sum\limits_{t = {t_{\rm{r}}}}^{{t_{{\rm{max}}}}} {{\rm{exp}}[\left( { - F{S_t}{A_t} - M} \right)(t - {t_{\rm{c}}})]} \\ \;\;\;\;\;{\rm{exp}}[ - M({t_{\rm{c}}} - {t_{\rm{r}}})]aL_t^b{G_t} \end{array} $ | (11) |
式中, SSB:总产卵亲体量, 单位为g;
SSBRF:捕捞死亡率(F)不为零时, 单位补充量产卵亲体量, 单位为g;
SSBRF=0:捕捞死亡率(F)为零时, 单位补充量产卵亲体量, 单位为g;
R:补充量, 假设为1;
F:捕捞死亡率;
M:自然死亡率;
a和b:体长与体重关系式参数, 引自公式(3)和公式(4);
Lt: t龄时的平均体长, 引自公式(1)和(2);
At: t龄时是否处于禁渔期, 如果处于其数值为0, 反之为1;
tmax:最大年龄, 单位为月份;
tr:补充年龄, 等于渔获物中的最小年龄, 单位为月份;
t:年龄, 单位为月份;
tc:起捕年龄, 单位为月份;
Gt: t龄时的成熟鱼类比例, 引自公式(5)和公式(6);
St: t龄时网具的选择系数, 由于缺乏网具选择系数方面的数据, 本研究假设网具对异齿裂腹鱼选择类型为“刀刃型”选择, 即达到起捕年龄tc, 选择系数值为1, 小于起捕年龄tc, 选择系数值为0。
(2) 单位补充量渔获量(YPR)计算公式如下:
| $ \begin{array}{l} {\rm{YPR}} = \frac{Y}{R} = \mathop \sum \limits_{t = {t_{\rm{r}}}}^{{t_{{\rm{max}}}}} \frac{{F{S_t}{A_t}}}{{F{S_t}{A_t} + M}}{\rm{exp}}( - {F_s}{S_t}{A_t} - M)(t - {t_{\rm{c}}}) \times \\ {\rm{exp}}[ - M({t_{\rm{c}}} - {t_{\rm{r}}})]{\rm{[}}1 - {\rm{exp(}} - F{S_t}{A_t} - M{\rm{)]}}aL_t^b \end{array} $ | (12) |
式中, Y:同一世代的总渔获量;
R:补充量, 假设为1;
F:捕捞死亡率;
M:自然死亡率;
a和b:体长与体重关系式参数, 引自公式(3)和公式(4);
Lt: t龄时的平均体长, 引自公式(1)和公式(2);
At: t龄时是否处于禁渔期, 如果处于其数值为0, 反之为1;
tmax:最大年龄, 单位为月份;
tr:补充年龄, 等于渔获物中的最小年龄, 单位为月份;
t:年龄, 单位为月份;
tc:起捕年龄, 单位为月份;
St: t龄时网具的选择系数, 由于缺乏网具选择系数方面的数据, 本研究假设网具对异齿裂腹鱼选择类型为“刀刃型”选择, 即达到起捕年龄tc, 选择系数值为1, 小于起捕年龄tc, 选择系数值为0。
对于已开发的鱼类种群, 准确地估算其自然死亡率是极其困难的。因此, 采用2个经验公式获取异齿裂腹鱼的自然死亡率区间, 并评估单位补充量模型对自然死亡率的敏感性。同时, 模拟14个不同的渔业养护措施评估起捕年龄和禁渔期对异齿裂腹鱼的保护效果(表 2)。
|
|
表 2 异齿裂腹鱼渔业养护措施 Tab.2 Conversation policies for Schizothorax oconnori in the Yarlung Zangbo River |
利用F25%和F40%两个参考点评价种群开发程度。F25%和F40%是最大单位补充量产卵亲体生物量的25%和40%所对应的捕捞死亡系数。F25%是下限参考点, 如果捕捞死亡系数高于该值, 说明种群被过度开发, 自然繁殖被严重破坏, 补充量不能维持种群的平衡稳定[20-22]。F40%是目标参考点, 是合理开发种群资源的捕捞标准, 如果捕捞死亡系数处于该值左右, 表明在保持种群稳定的前提下可以提供高产量[23]。
2 结果与分析 2.1 死亡参数Chapman-Robson法估算的雌鱼和雄鱼的总瞬时死亡率(Z)分别为0.11/a和0.16/a。极限年龄法估算的雌鱼和雄鱼的自然死亡率(M)分别为0.08/a和0.10/a; 而生长方程参数法估算的自然死亡率(M)则分别为0.09/a和0.12/a。因此, 雌鱼的自然死亡率(M)范围为0.08/a~0.09/a, 而雄鱼的自然死亡率(M)范围为0.10/a~0.12/a。对应的雌鱼的当前捕捞死亡率(Fcur)范围为0.02/a~0.03/a, 而雄鱼的当前捕捞死亡率(Fcur)范围为0.04/a~0.06/a。
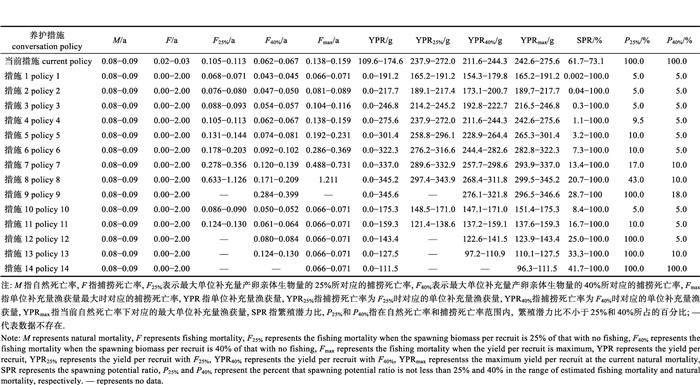
2.2 资源状况在现有的渔业养护措施下, 采用单位补充量模型来分析异齿裂腹鱼雌性和雄性种群的资源现状对自然死亡率的敏感性。在估算的自然死亡率范围内, 雌鱼种群的繁殖潜力比为61.7%~73.1%, 全部高于40%, 雌鱼种群的当前捕捞死亡率为0.02/a~0.03/a, 全部低于目标参考点(F40%); 雄鱼种群的繁殖潜力比为48.5%~63.3%, 全部高于40%, 雄鱼种群的当前捕捞死亡率为0.04/a~0.06/a, 全部低于目标参考点(F40%)。单位补充量渔获量分析表明, 雌鱼种群在捕捞死亡率为0.138/a~0.159/a时, 其单位补充量渔获量达到最大值; 而雄鱼种群在捕捞死亡率为0.178/a~0.228/a时, 其单位补充量渔获量达到最大值(表 3和表 4)。上述结果表明, 在现有的渔业养护措施下, 异齿裂腹鱼种群的利用基本合理, 但要防止长期持续利用对种群的不利影响。
|
|
表 3 养护措施对异齿裂腹鱼雌性种群的生物学参考点、单位补充量渔获量和繁殖潜力比的影响 Tab.3 The effects of different conversation policies on biological reference points, yield perrecruit and spawning potential ratio (SPR) of female Schizothorax oconnori |
|
|
表 4 养护措施对异齿裂腹鱼雄性种群的生物学参考点、单位补充量渔获量和繁殖潜力比的影响 Tab.4 The effects of different conversation policies on biological reference points, yield per recruit and spawning potential ratio (SPR) of male Schizothorax oconnori |
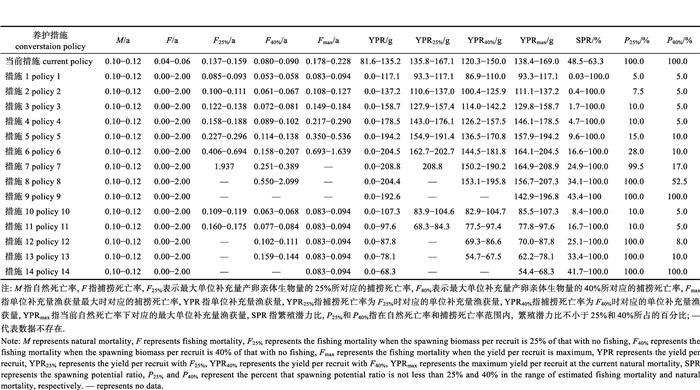
在设定的自然死亡率和捕捞死亡率值域内, 利用单位补充量模型评估了起捕年龄对异齿裂腹鱼种群资源的影响(表 3和表 4)。随着起捕年龄的逐渐增大, 异齿裂腹鱼雌性种群的YPR、YPRmax、SPR、P25%和P40%表现为逐渐增大的趋势, 而异齿裂腹鱼雄性种群的YPR和YPRmax表现为先增大后减小的趋势, SPR、P25%和P40%表现为逐渐增大的趋势, 这说明对于异齿裂腹鱼种群来说, 提高起捕年龄是一种有效的资源养护策略。因此, 我们期望利用YPR和SPR等值线图来确定最优的起捕年龄。
在起捕年龄的大部分值域内, 单位补充量渔获量增速是随着捕捞强度的增大逐渐减缓的(图 3)。在设定的自然死亡率值域内, 与当前单位补充量渔获量相比, 将异齿裂腹鱼类雌性和雄性种群的起捕年龄分别设置为8~17龄和6~13龄, 其单位补充量渔获量在捕捞死亡率大部分值域上波动幅度相对较小(图 3)。此外, 在设定的自然死亡率和捕捞死亡率值域内, 将异齿裂腹鱼雌性和雄性种群的起捕年龄分别提高至不小于17龄和14龄, 能够保证其繁殖潜力比始终高于下限参考点(25%); 而将雌性和雄性的起捕年龄分别提高至不小于19龄和17龄, 能够保证其繁殖潜力比始终高于目标参考点(40%, 图 4)。结合当前异齿裂腹鱼资源利用基本合理的现状, 在牺牲单位补充量渔获量的基础上, 建议将异齿裂腹鱼的起捕年龄设置为不小于17龄, 以防止长期持续利用对其资源造成的不利影响。

|
图 3 雅鲁藏布江异齿裂腹鱼单位补充量渔获量等值线图
红点代表估算的当前单位补充量渔获量; 图中自然死亡率(M)分别为雌鱼和雄鱼估算的自然死亡率范围的最小值、中值和最大值.
Fig.3 Isopleths of yield per recruit for Schizothorax oconnori in the Yarlung Zangbo River
Red points represent the current estimated yield per recruit. The natural mortalities in the figure are minimum values, media values and maxinmum values of the estimated natural mortality ranges for female and male, respectively. |

|
图 4 雅鲁藏布江异齿裂腹鱼繁殖潜力比等值线图
红点代表估算的当前繁殖潜力比; 图中自然死亡率(M)分别为雌鱼和雄鱼估算的自然死亡率范围的最小值、中值和最大值.
Fig.4 Isopleths of spawning potential ratio for Schizothorax oconnori in the Yarlung Zangbo River
Red points represent the current estimated spawning potential ratio. The natural mortalities in the figure are minimum values, media values and maximum values of the estimated natural mortality ranges for female and male, respectively. |
在设定的自然死亡率和捕捞死亡率值域内, 禁渔期导致单位补充量渔获量降低, 但能够对异齿裂腹鱼种群资源形成有效的保护(表 3和表 4)。随着禁渔期的增加, 异齿裂腹鱼雌性和雄性种群的单位补充量渔获量呈现持续下降趋势, 其繁殖潜力比却呈现持续上升趋势。在设定的自然死亡率和捕捞死亡率值域内, 将禁渔时间设置为2—4月, 异齿裂腹鱼雌性和雄性种群的繁殖潜力比范围分别为25.0%~100.0%和25.1%~ 100.0%, 将禁渔时间设置为2—5月, 其繁殖潜力比范围分别为33.3%~100.0%和33.4%~100.0%, 将禁渔时间设置为2—6月, 其雌雄鱼繁殖潜力比范围都为41.7%~100.0%。因此, 建议将禁渔期设置为至少2—5月, 以防止长期持续利用对异齿裂腹鱼种群资源造成的不利影响。
3 讨论自然死亡率是开展种群动态评估的重要参数之一。对于已开发的自然种群, 自然死亡率通常与捕捞死亡率交织在一起, 因此, 准确地估算自然死亡率是极其困难的[15, 24-25]。估算自然死亡率通常采用直接估算法和经验公式法(间接估算法)。直接估算法常常需要大数据的支持, 其仅能运用于研究数据相对较为丰富的种群; 为了解决研究数据相对匮乏种群的自然死亡率估算问题, 在过去的70多年中, 学者们利用鱼类的生活史信息构建了许多经验公式来估算自然死亡率[15-16, 26-28]。与直接估算法相比, 由于经验公式法所利用的数据较少, 其估算结果的可靠性必然相对较低, 因此, 在种群动态研究过程中, 学者们通常采用至少2种经验公式来评估自然死亡率的不确定性。本研究结果表明, 在估算的自然死亡率范围内, 异齿裂腹鱼资源现状对自然死亡率的敏感性较差, 即自然死亡率的变化无法引起其资源开发现状的改变, Smith等[14]报道采用Chapman-Robson法估算的总死亡率常常产生负偏差, 这暗示本研究估算的当前捕捞死亡率偏低, 此外, 本研究利用的样本为10年前采集, 其时效性稍显不足。综上所述, 在评估栖息于雅鲁藏布江中游异齿裂腹鱼资源利用现状时需保持保守且偏悲观的结果。
繁殖期实施禁渔通常在3种情况下能够对目标种群产生有效的保护: (1)捕捞对鱼类的繁殖活动产生了干扰; (2)目标种群在产卵场集群从而使其更容易捕获; (3)禁渔期能够减少周年的捕捞努力量[29]。研究结果表明, 在繁殖期实施禁渔能够有效地保护异齿裂腹鱼种群资源, 其保护效果随着禁渔期的延长而增加。马宝珊[6]研究结果表明, 异齿裂腹鱼是典型的春季产卵鱼类, 具有集群产卵的繁殖特性。此外, 野外调查发现, 在繁殖季节渔获物中大型个体的比例显著性增加暗示集群产卵可能增大了异齿裂腹鱼的可捕性。因此, 繁殖季节实施禁渔是一种行之有效的保护策略。
渔业资源养护是一个庞杂的系统工作, 其可能涉及渔业法规、风俗习惯、地理环境、资源现状、社会经济等诸多方面。根据异齿裂腹鱼的生物学特性以及本文的研究结果, 建议从以下几方面着手来养护其资源。
(1) 完善西藏本土的渔业法规。以《中华人民共和国渔业法》和《中华人民共和国野生动物保护法》等法律法规为依据, 结合自治区地理环境、风俗习惯以及渔业资源具体情况, 因地制宜地修改现有地方渔业法规, 积极探索主动式渔业资源养护模式, 形成完善的渔业法规体系。2006年颁布实施的《西藏自治区实施〈中华人民共和国渔业法〉办法》中明确提出异齿裂腹鱼的最低起捕体重为400 g。基于本文研究结果, 建议将异齿裂腹鱼的最低起捕体重修改为1250 g。同时, 建议将渔业资源养护和渔业法规与制度的考核作为捕捞许可证发放或者审核的必备条件。
(2) 加强渔政执法队伍和设备建设。健全的地方渔业法规体系对渔业资源养护固然重要, 但如果没有强大的监督执法力量来实现渔业资源有效养护, 再健全的渔业法规体系只会沦落为一纸空文。虽然《西藏自治区实施〈中华人民共和国渔业法〉办法》早在13年前已经颁布并实施, 但自治区主要水生生物资源的衰退趋势始终未能得到有效遏止, 主要原因之一是监督执法力度始终较弱。建议在现有三级渔政管理机构的基础上, 进一步完善渔业综合执法管理体系建设, 增加具有水产专业背景管理人员编制, 对执法管理人员定期进行相关法律法规和专业知识的培训, 提高执法水平。同时应当进一步提升渔业执法装备水平, 例如, 为执法人员配备执法交通工具和取证仪器等, 提高执法能力。
(3) 建议在雅鲁藏布江的中游水域实施全面禁渔。尽管提高起捕年龄和实施禁渔期两种方法都能够有效地保护提高起捕年龄, 但禁渔期相对更容易实施和监管, 这种情况在地广人稀以及渔政监管人员匮乏的地区表现尤为突出。野外调查发现, 藏族同胞有放生的风俗习惯, 其喜好购买小型鱼类以增加放生量。当地渔民的经济收入可被划分为两部分, 一部分来自于出售成鱼的收入, 另一部分来自于出售小型放生鱼类收入。如果通过提高起捕年龄来保护异齿裂腹鱼的资源, 当地渔民依赖于小型放生鱼类的收入必定减少; 同理如果通过实施禁渔期来保护异齿裂腹鱼的资源, 当地渔民依赖于成鱼的收入也必定减少。因此, 无论采用何种养护措施, 都会造成当地渔民经济收入的降低。提高起捕年龄来保护异齿裂腹鱼的资源, 必然导致土著小型鱼类的渔获量大幅缩减, 最终引起藏族同胞购买外来小型鱼类(如泥鳅和鲫鱼等)进行放生, 从而加剧外来鱼类对裂腹鱼类预备群体和补充群体的威胁。尽管如此, 禁渔期过后, 那些付费取得捕捞许可证的渔民自然就会增加捕鱼, 弥补禁渔期给他们造成的经济损失, 这必然造成禁渔效果大打折扣。通过借鉴长江流域的禁渔经验, 结合异齿裂腹鱼雌性的初次性成熟年龄为9.5龄, 故建议在雅鲁藏布江的中游水域实施至少10年的全面禁渔。
(4) 建议实施生态补偿。由于西藏特殊的宗教信仰, 藏族同胞珍爱生命, 反对杀生, 几乎不从事捕捞业。雅鲁藏布江流域的渔民绝大多数是来自四川等地的渔民, 通过引导渔民转产转业, 禁渔不会影响他们的经济收入, 且有利于促进和谐社会的建设。此外, 通过完善土著鱼类增殖放流技术规范等措施提高异齿裂腹鱼的增殖放流效果, 并将藏族同胞的放生纳入人工放流总体安排, 例如, 向藏族同胞免费提供土著鱼类仔稚鱼, 既可满足放生愿望, 达到增殖放流目的, 又可控制外来鱼类入侵。
(5) 实施水电工程影响的减缓措施。雅鲁藏布江蕴藏着丰富的水能资源, 全流域水能蕴藏量超过1.13×108 kW。雅鲁藏布江水资源的合理开发利用, 对于西藏地区乃至全国的国民经济可持续发展具有深远意义[30]。雅鲁藏布江已制定了水电开发规划, 已建成和正在修建的有藏木、加查、直孔、旁多、多布、老虎嘴、雪卡等多座水电站。水电工程的建设改变了天然河道的形态、水文情势和水体理化性质等, 对异齿裂腹鱼种群及其赖以生存的水生态系统造成影响。建议通过加强对水利工程的监管和审批的力度、保护异齿裂腹鱼的天然生境、加强异齿裂腹鱼的增殖放流管理以及设立水域生态和鱼类资源保护专项基金等途径减缓水电工程对异齿裂腹鱼种群资源的不利影响。
(6) 突破异齿裂腹鱼增养殖关键技术。近十几年来, 随着当地社会经济的发展和人们消费观念的改变, 人们对水产品, 尤其是本地特色水产品的需求日益增长, 从自然水体中捕捞的异齿裂腹鱼除了满足本地的消费需求外, 大量鲜鱼销往重庆和成都等全国各大城市。市场需求刺激了对自然种群的掠夺式开发, 建议通过人工养殖满足市场需求, 减少对自然种群的捕捞强度, 对加强异齿裂腹鱼资源保护具有极其重要的意义。
4 结论异齿裂腹鱼属于裂腹鱼类, 随着青藏高原的隆起, 其由第三纪晚期的原始鲃亚科鱼类逐渐进化而来[4, 31-32]。异齿裂腹鱼是当地水域生态系统初级消费者的重要组成, 其资源量的变动、消失或者迁徙将通过生态级联效应引起水域生态系统结构和功能的失衡, 最终给区域经济和社会的可持续发展带来严重威胁。因此, 掌握异齿裂腹鱼的种群动态数据是科学合理地开展其资源养护乃至维系雅鲁藏布江水域生态系统结构和功能完整性的重要基础。本研究结果表明, 在现有的渔业养护措施下, 异齿裂腹鱼种群的利用基本合理, 但要防止长期持续利用对种群的不利影响; 综合考虑西藏当地的地理环境、风俗习惯、异齿裂腹鱼自身的生物特性以及不同养护措施对异齿裂腹鱼资源的保护效果, 本研究建议综合采用完善渔业法规、加强渔业执法力度、实施全面禁渔、实施生态补偿和水电工程影响的减缓措施以及突破异齿裂腹鱼增养殖技术等措施来养护异齿裂腹鱼种群资源。
| [1] |
Cao W X. The ecological protection of the water area in the hydropower cascade development of the upper reaches of the Yangtze river[J]. Yangtze River Technical Economy, 2017, 1(1): 25-30. [曹文宣. 长江上游水电梯级开发的水域生态保护问题[J]. 长江技术经济, 2017, 1(1): 25-30.] |
| [2] |
Xiong F, Liu H Y, Duan X B, et al. Population parameters and population abundance of Rhinogobio cylindricus in Zhuyangxi section of the upper Yangtze River[J]. Acta Ecologica Sinica, 2015, 35(22): 7320-7327. [熊飞, 刘红艳, 段辛斌, 等. 长江上游朱杨溪江段圆筒吻鮈种群参数和资源量[J]. 生态学报, 2015, 35(22): 7320-7327.] |
| [3] |
Xiong F, Liu H Y, Duan X B, et al. Estimating population abundance and utilization of Rhinogobio ventralis, an endemic fish species in the upper Yangtze River[J]. Biodiversity Science, 2016, 24(3): 304-312. [熊飞, 刘红艳, 段辛斌, 等. 长江上游特有种长鳍吻鮈种群数量和资源利用评估[J]. 生物多样性, 2016, 24(3): 304-312.] |
| [4] |
Cao W X, Chen Y Y, Wu Y F, et al. Origin and evolution of Schizothoracine fishes in relation to the upheaval of the Xizang Plateau[C]//Tibetan Expedition Team of the Chinese Academy of Science Studies. On the Period, Amplitude and Type of the Uplift of the Qinghai-Xizang Plateau. Beijing: Science Press, 1981: 118-130. [曹文宣, 陈宜瑜, 武云飞, 等.朱松泉裂腹鱼类的起源和演化及其与青藏高原隆起的关系[C]//中国科学院青藏高原综合科学考察队.青藏高原隆起的时代、幅度和形式问题.北京: 科学出版社, 1981: 118-130.]
|
| [5] |
Wu Y F, Wu C Z. Fish of Qinghai-Tibet Plateau[M]. Chengdu: Sichuan Science and Technology Press, 1992: 1-599. [武云飞, 吴翠珍. 青藏高原鱼类[M]. 成都: 四川科学技术出版社, 1992: 1-599.]
|
| [6] |
Ma B S. Study on the biology and population dynamics of Schizothorax o'connori[D]. Wuhan: Huazhong Agricultural University, 2011: 1-154. [马宝珊.异齿裂腹鱼个体生物学和种群动态研究[D].武汉: 华中农业大学, 2011: 1-154.]
|
| [7] |
He Z T. Studies on age and growth of Schizothorax o'connori in Lhasa River in Tibet[D]. Wuhan: Huazhong Agricultural University, 2005: 1-69. [贺舟挺.西藏拉萨河异齿裂腹鱼年龄与生长的研究[D].武汉: 华中农业大学, 2005: 1-69.]
|
| [8] |
Yao J L, Chen Y F, Chen F, et al. Age and growth of an endemic Tibetan fish, Schizothorax o'connori, in the Yarlung Tsangpo River[J]. Journal of Freshwater Ecology, 2009, 24(2): 343-345. DOI:10.1080/02705060.2009.9664303 |
| [9] |
Chen F, Chen Y F. Investigation and protection strategies of fishes of Lhasa River[J]. Acta Hydrobiologica Sinica, 2010, 34(2): 278-285. [陈锋, 陈毅峰. 拉萨河鱼类调查及保护[J]. 水生生物学报, 2010, 34(2): 278-285.] |
| [10] |
Liu H P, Mou Z B, Cai B, et al. Coupling supply-side structural reform and technological innovation boosting the fishery resource conservation process in Tibet[J]. Journal of Lake Sciences, 2018, 30(1): 266-278. [刘海平, 牟振波, 蔡斌, 等. 供给侧改革与科技创新耦合助推西藏渔业资源养护[J]. 湖泊科学, 2018, 30(1): 266-278.] |
| [11] |
Yang H Y, Huang D M, Xie S, et al. Status quo of fishery resources in the middle reach of Brahmaputra River[J]. Journal of Hydroecology, 2010, 31(6): 120-126. [杨汉运, 黄道明, 谢山, 等. 雅鲁藏布江中游渔业资源现状研究[J]. 水生态学杂志, 2010, 31(6): 120-126.] |
| [12] |
Fan L Q, Liu H P, Lin J, et al. Non-native fishes: distribution and assemblage structure in the Lhasa River Basin, Tibet, China[J]. Acta Hydrobiologica Sinica, 2016, 40(5): 958-967. [范丽卿, 刘海平, 林进, 等. 拉萨河流域外来鱼类的分布、群落结构及其与环境的关系[J]. 水生生物学报, 2016, 40(5): 958-967.] |
| [13] |
Li J C, Tu Y L, Lin J, et al. Influence of religious release on fish assemblages in the Lhasa River Basin, Tibet, China[J]. Journal of Plateau Agriculture, 2018, 2(5): 462-469. [李建川, 土艳丽, 林进, 等. 放生对拉萨河流域鱼类物种多样性和群落结构的影响[J]. 高原农业, 2018, 2(5): 462-469.] |
| [14] |
Smith M W, Then A Y, Wor C, et al. Recommendations for catch-curve analysis[J]. North American Journal of Fisheries Management, 2012, 32(5): 956-967. DOI:10.1080/02755947.2012.711270 |
| [15] |
Then A Y, Hoenig J M, Hall N G, et al. Evaluating the predictive performance of empirical estimators of natural mortality rate using information on over 200 fish species[J]. ICES Journal of Marine Science, 2015, 72(1): 82-92. DOI:10.1093/icesjms/fsu136 |
| [16] |
Hoenig J M. Empirical use of longevity data to estimate mortality rates[J]. Fishery Bulletin, 1983, 82(1): 898-903. |
| [17] |
Taylor C C. Cod growth and temperature[J]. ICES Journal of Marine Science, 1958, 23(3): 366-370. DOI:10.1093/icesjms/23.3.366 |
| [18] |
Goodyear C P. Spawning stock biomass per recruit in fisheries management: Foundation and current use[J]. Canadian Special Publication of Fisheries and Aquatic Sciences, 1993, 120: 67-81. |
| [19] |
Quinn Ⅱ T J, Deriso R B. Quantitative Fish Dynamics[M]. New York: Oxford University Press, 1999: 1-542.
|
| [20] |
Griffiths M H. The application of per-recruit models to Argyrosomus inodorus, an important South African sciaenid fish[J]. Fisheries Research, 1997, 30(1-2): 103-115. DOI:10.1016/S0165-7836(96)00552-8 |
| [21] |
Kirchner C H. Fisheries regulations based on yield-per-recruit analysis for the linefish silver kob Argyrosomus inodorus in Namibian waters[J]. Fisheries Research, 2001, 52(3): 155-167. |
| [22] |
Sun C L, Ehrhardt N M, Porch C E, et al. Analyses of yield and spawning stock biomass per recruit for the South Atlantic albacore (Thunnus alalunga)[J]. Fisheries Research, 2002, 56(2): 193-204. |
| [23] |
Sun C L, Wang S P, Porch C E, et al. Sex-specific yield per recruit and spawning stock biomass per recruit for the swordfish, Xiphias gladius, in the waters around Taiwan[J]. Fisheries Research, 2005, 71(1): 61-69. |
| [24] |
Lee H H, Maunder M N, Piner K R, et al. Estimating natural mortality within a fisheries stock assessment model: An evaluation using simulation analysis based on twelve stock assessments[J]. Fisheries Research, 2011, 109(1): 89-94. |
| [25] |
Kenchington T J. Natural mortality estimators for information-limited fisheries[J]. Fish and Fisheries, 2014, 15(4): 533-562. DOI:10.1111/faf.12027 |
| [26] |
Beverton R J H, Holt S J. A review of the lifespans and mortality rates of fish in nature, and their relation to growth and other physiological characteristics[M]//Wolstenholme G E W, O'Conner M. Ciba Foundation Symposium—The Lifespan of Animals (Colloquia on Ageing). Chichester: John Wiley and Sons, 1959: 142-180.
|
| [27] |
Pauly D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks[J]. ICES Journal of Marine Science, 1980, 39(2): 175-192. DOI:10.1093/icesjms/39.2.175 |
| [28] |
Jensen A L. Beverton and Holt life history invariants result from optimal trade-off of reproduction and survival[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1996, 53(4): 820-822. DOI:10.1139/f95-233 |
| [29] |
Arendse C J, Govender A, Branch G M. Are closed fishing seasons an effective means of increasing reproductive output? A per-recruit simulation using the limpet Cymbula granatina as a case history[J]. Fisheries Research, 2007, 85(1-2): 93-100. DOI:10.1016/j.fishres.2007.01.001 |
| [30] |
Xu D M, Chen C Y, Liang W Y. Hydropower development at the Yalu Tsangpo River[J]. Engineering Science, 2002, 4(12): 47-52. [徐大懋, 陈传友, 梁维燕. 雅鲁藏布江水能开发[J]. 中国工程科学, 2002, 4(12): 47-52. DOI:10.3969/j.issn.1009-1742.2002.12.009] |
| [31] |
He D K, Chen Y F. Biogeography and molecular phylogeny of the genus Schizothorax (Teleostei: Cyprinidae) in China inferred from cytochrome b sequences[J]. Journal of Biogeography, 2006, 33(8): 1448-1460. DOI:10.1111/j.1365-2699.2006.01510.x |
| [32] |
He D K, Chen Y F. Molecular phylogeny and biogeography of highly specialized schizothoracine fishes[J]. Chinese Science Bulletin, 2007, 52(3): 303-312. [何德奎, 陈毅峰. 高度特化等级裂腹鱼类分子系统发育与生物地理学[J]. 科学通报, 2007, 52(3): 303-312. DOI:10.3321/j.issn:0023-074X.2007.03.010] |